细胞呼吸与能量转换
绿皮和蓝皮细胞生物学在细胞呼吸这一节都有许多的错误,生物化学与分子生物学中也没有特别明确地讲明几个重要问题。
在细胞内特定的细胞器(主要是线粒体)内,在O2的参与下,分解各种大分子物质,产生CO2;与此同时,分解代谢所释放出的能量储存于ATP中,这一过程称为细胞呼吸(cellular respi- ration),也称为生物氧化(biological oxidation)或细胞氧化(cellular oxidation)。
细胞呼吸是细胞内提供生物能源的主要途径,它的化学本质与燃烧反应相同,最终产物都是CO2和H2O,释放的能量也相等。但是,细胞呼吸具有以下特点:
- 本质上是在线粒体中进行的一系列由酶系所催化的氧化还原反应;
- 产生的能量储存于ATP的高能磷酸键中;
- 整个反应过程是分步进行的,能量也是逐步释放的;
- 反应是在恒温(37℃)和恒压条件下进行的;
- 反应过程中需要H20的参与。
ATP的放能、储能反应简式如下:
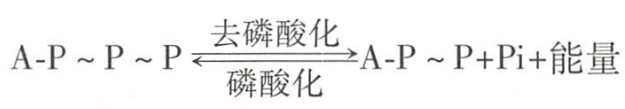
ATP是“能量货币”。ATP是细胞生命活动的直接供能者,也是细胞内能量获得、转换、储存和利用等环节的联系纽带。
ATP中所携带的能量来源于糖、氨基酸和脂肪酸等的氧化。
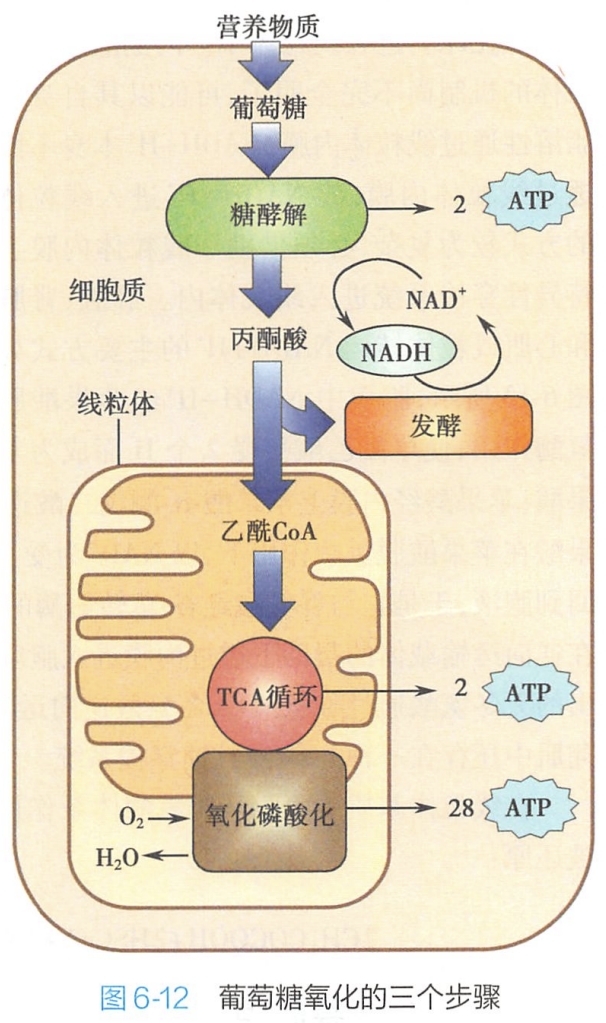
- 以葡萄糖氧化为例,从糖酵解到ATP的形成是一个极其复杂的过程,大体分为三个步骤:即糖酵解(glycolysis)、三羧酸循环(tricarboxylic acid cycle,TCA cycle)和氧化磷酸化(oxidative phosphorylation)(图6-12)。
- 蛋白质和脂肪的彻底氧化只在第一步中与糖有所区别。
一、葡萄糖在细胞质中的糖酵解
(一)糖酵解有三种代谢去路
糖酵解在细胞质中进行,其过程可概括为以下方程式:
1(葡萄糖)→2:2×1:0:(2×2-2)(丙酮酸:NADH+H+:FADH2:ATP)两次底物磷酸化
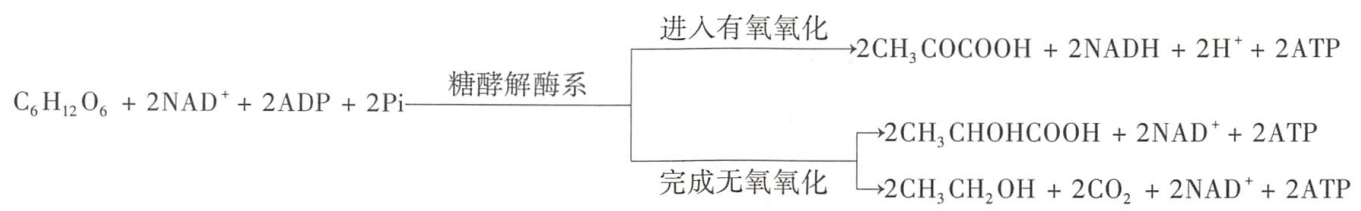
- 1分子葡萄糖经过十多步反应,生成2分子丙酮酸,同时脱下2对H形成 2分子NADH+H+。NAD+能可逆地接受2个电子和1个H+,另1个H+则留在溶质中。
- 在糖酵解过程中一共生成4分子ATP,但由于要消耗2分子ATP,所以净生成2分子的ATP。若从糖原开始糖酵解,因不需消耗1分子ATP使葡萄糖磷酸化,则总反应净生成3分子ATP。
- 这种由高能底物水解放能,直接将高能磷酸键从底物转移到ADP上,使ADP磷酸化生成ATP的作用,称为底物水平磷酸化(sub-strate-level phosphorylation)。
糖酵解产物丙酮酸的代谢去路,因不同生活状态的生物而异。专性厌氧生物在无氧情况下,丙酮酸可由NADH+H+供氢而还原为乳酸或乙醇,从而完成无氧氧化过程。
(二)丙酮酸与NADH+H+进入线粒体
专性需氧生物在供氧充足时,丙酮酸与NADH+H+将作为有氧氧化原料进入线粒体中。
- 丙酮酸进入线粒体的机制尚不完全明了,可能以其自身的脂溶性通过线粒体内膜;
- NADH+H+本身不能透过线粒体内膜,必须借助于线粒体内膜上特异性穿梭系统进入线粒体内。
肝脏、肾脏和心肌线粒体转运NADH+H+的主要方式如图6-13所示
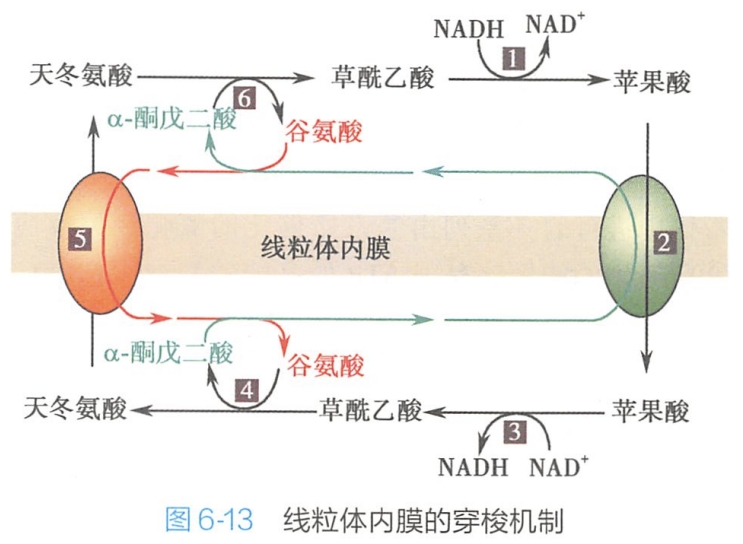
- 胞质中NADH+H+经苹果酸脱氢酶作用,使草酰乙酸接受2个H而成为苹果酸;苹果酸经内膜上苹果酸-Q-酮戊二酸逆向运输载体的变构作用转入线粒体内;
- 进入线粒体的苹果酸在苹果酸脱氢酶作用下,以NAD+为受氢体形成草酰乙酸和NADH+H+;
- 而草酰乙酸不能经内膜回到胞液,于是它与谷氨酸经谷-草转氨酶的作用而相互转变为天冬氨酸和α-酮戊二酸,这两者都能在逆向运输载体的帮助下透过内膜进入胞质中去;
- 线粒体内消耗的谷氨酸则由胞液内的谷氨酸与外出的天冬氨酸通过谷氨酸-天冬氨酸逆向运输载体实现交换运输以取得补充。
另外,在脑和昆虫的飞翔肌中还存在一种α-磷酸甘油穿梭系统。 #生化
在线粒体基质中丙酮酸脱氢酶体系作用下,丙酮酸进一步分解为乙酰辅酶A,NAD+作为受氢体被还原:
1(葡萄糖)→2:2×1:0:(2×2-2)(丙酮酸:NADH+H+:FADH2:ATP)
2(丙酮酸)→2(1:1:0:0)看脱碳或去氢,CH2
2CH3COCOOH+2HSCoA+2NAD+→2CH3COScoA+2CO2+2NADH+2H+
二、线粒体基质中的三羧酸循环
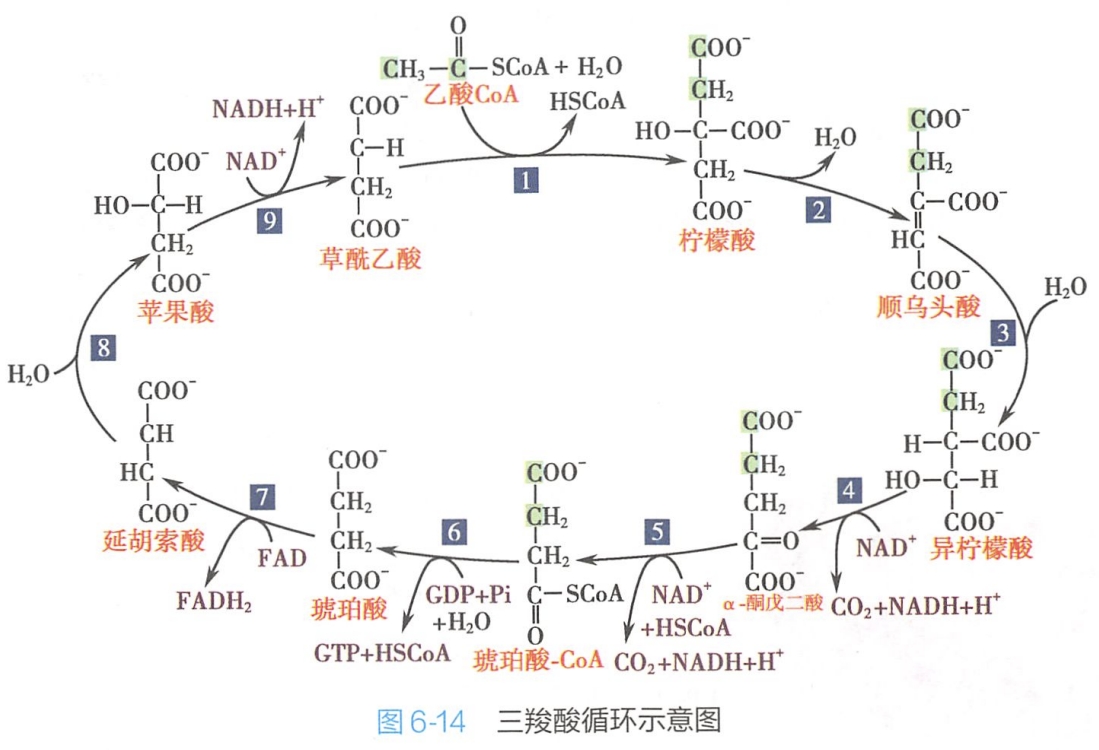
在线粒体基质中,乙酰CoA与草酰乙酸结合成柠檬酸而进入柠檬酸循环,由于柠檬酸有3个羧基,故也叫三羧酸循环(TCA循环)(图6-14)。
循环中,柠檬酸经过一系列酶促的氧化脱氢和脱羧反应,其中的2个碳原子氧化形成CO2,从而削减了2个碳原子。整个过程中,总共消耗了3分子H20,生成1分子的GTP(可转换为1分子的ATP)、4对H和2分子CO2。脱下的4对H,其中3对以NAD+为受氢体,另1对以FAD为受氢体。FAD能可逆地接受2个H,即2个质子和2个电子,转变成还原态FADH2。
三羧酸循环总的反应式为:
1(葡萄糖)→2:2×1:0:(2×2-2)(丙酮酸:NADH+H+:FADH2:ATP)
2(丙酮酸)→2(1:1:0:0)(乙酰CoA:NADH+H+:FADH2:ATP)看脱碳
2(1:3)(乙酰CoA:水)→2(2:3:1:1)(CO2:NADH+H+:FADH2:ATP)看少氢
2CH3COSCoA+6NAD++2FAD+2GDP+2Pi+6H2O→4CO2+6NADH+6H++2FADH2+2HSCoA+2GTP
(书上公式和上图都有错误,太离谱)
- 三羧酸循环是各类有机物相互转化的枢纽。三羧酸循环的中间产物可用来合成包括氨基酸、卟啉及嘧啶核苷酸在内的许多物质。
- 三羧酸循环是各种有机物进行最后氧化的过程。除了丙酮酸外,脂肪酸和一些氨基酸也从细胞质进入线粒体,并进一步转化成乙酰CoA或三羧酸循环的其他中间体。只有经过三羧酸循环,有机物才能进行完全氧化,提供远比糖无氧酵解所能提供的多得多的能量,供生命活动的需要。
- ATP/ADP及NADH/NAD+比值高时均能降低三羧酸循环的速度。
三、氧化磷酸化耦联
氧化磷酸化是释放代谢能的主要环节,在这个过程中,NADH和FADH2分子把它们从食物氧化得来的电子转移到氧分子。
这一反应相当于氢原子在空气中燃烧最终形成水的过程,释放出的能量绝大部分用于生成ATP,少部分以热的形式释放。
(一)呼吸链和ATP合酶复合体是氧化磷酸化的结构基础
1.呼吸链是一系列能够可逆地接受和释放H+和e-的酶体系
1分子的葡萄糖经无氧氧化、丙酮酸脱氢和三羧酸循环,共产生了6分子的CO2和12对H,这些H必须进一步氧化成为水,整个有氧氧化过程才告结束。
但H并不能与O2直接结合,一般认为H须首先离解为H+和e-,电子经过线粒体内膜上酶体系的逐级传递,最终使1/2O2,成为O2-,后者再与基质中的2个H+化合生成H20。
这一传递电子的酶体系是由一系列能够可逆地接受和释放H+和e-的化学物质所组成,它们在内膜上有序地排列成相互关联的链状,称为呼吸链(respiratory chain)或电子传递呼吸链(electron transport respiratory chain)。
氧化还原电位从低到高(氧化还原电位越低,越容易被氧化,失电子,越高则越容易得电子),电子转移导致自由能释放供质子泵(Ⅰ、Ⅲ、Ⅳ)泵H+到膜间腔
第一条链:NADH+H+、Ⅰ(FMN、FeS)、CoQ、Ⅲ(Cytb、FeS、Cytc1)、Cytc、Ⅳ(CuA、Cyta、Cyta3-CuB)、O2
第二条链:琥珀酸/其他、Ⅱ(FAD、FeS)、CoQ、Ⅲ(Cytb、FeS、Cytc1)、Cytc、Ⅳ(CuA、Cyta、Cyta3-CuB)、O2
只传递电子的酶和辅酶称为电子传递体,它们可分为醌类、细胞色素和铁硫蛋白三类化合物:
- 泛醌(辅酶Q,CoQ),CoQ是脂溶性的蛋白质,可在脂双层中从膜的一侧向另一侧移动; #? 不太可能
- 细胞色素c(Cytc)等。细胞色素c是膜周边蛋白,可在膜表面移动。
- 硫铁蛋白FeS
既传递电子又传递质子的酶和辅酶称为递氢体。
除了泛醌和细胞色素c之外,呼吸链其他成员分别组成了Ⅰ(NADH脱氢酶)、Ⅱ(琥珀酸脱氢酶,即三羧酸循环中那个)、Ⅲ、Ⅳ(细胞色素c氧化酶)四个脂类蛋白质复合体,它们是线粒体内膜的整合蛋白(表6-3)。
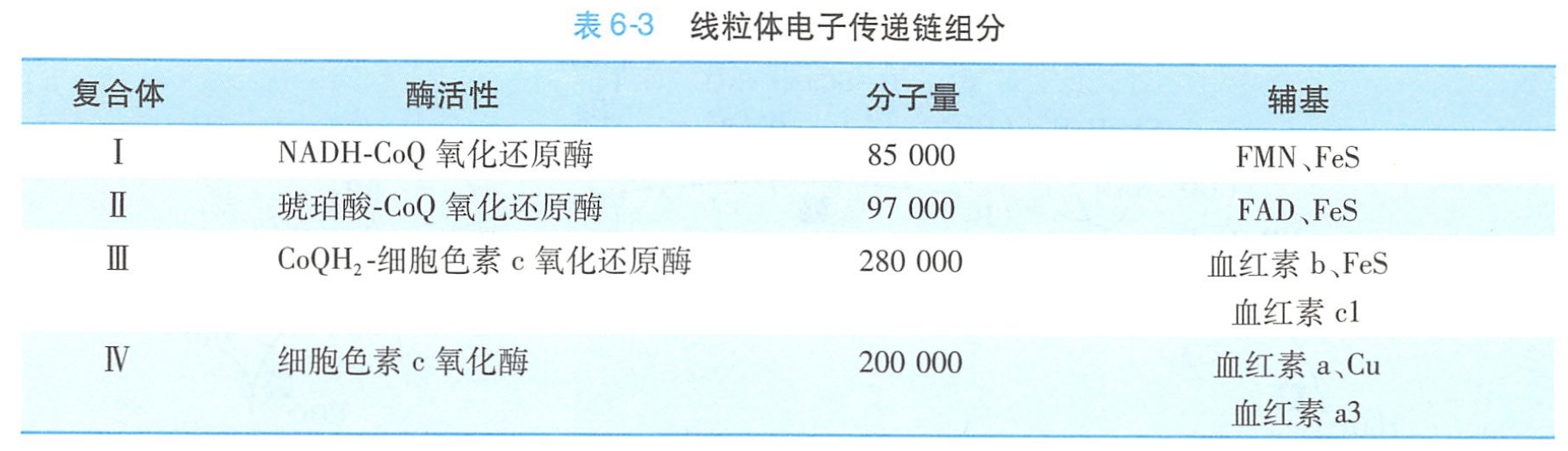
2.ATP合酶复合体催化ATP的合成
线粒体内膜(包括嵴)的内表面附有许多圆球形基粒。基粒由头部、柄部和基片3部分组成: 头部与柄部相连凸出在内膜表面,柄部则与嵌入内膜的基片相连。
基粒是将呼吸链电子传递过程中所释放的能量(质子浓度梯度和电位差)用于使ADP磷酸化生成ATP的关键装置,是由多种多肽构成的复合体,其化学本质是ATP合酶或ATP合酶复合体,也称F0F1ATP合酶(图6-15)。
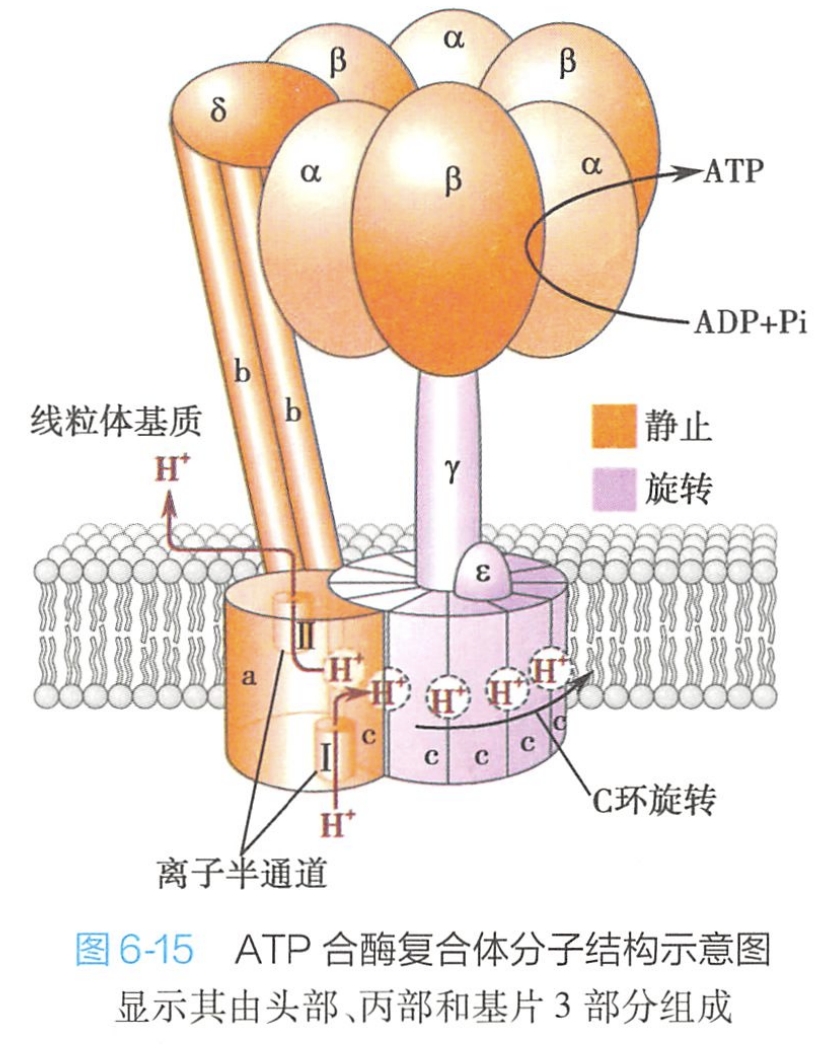
(1)头部:具有酶活性,又称耦联因子F1,是由五种亚基组成的α3β3γδε多亚基复合体。纯化的F1可催化ATP水解,但其在自然状态下(通过柄部与基片相连)的功能是催化ATP合成。F1因子可被F1抑制蛋白(F1 inhibitory protein)结合从而抑制ATP的合成。
(2)柄部:其实应该是F0的c亚基。连接头部与基片,这是一种对寡霉素敏感的蛋白质(OSCP)。OSCP能与寡霉素特异结合并使寡霉素的解耦联作用得以发挥,从而抑制ATP合成。
(3)基片:又称耦联因子F0。F0镶嵌于内膜的脂双层中,不仅起连接F1与内膜的作用,而且还是质子流向F1的穿膜通道。
(二)氧化磷酸化耦联
经糖酵解和三羧酸循环产生的NADH和FADH2是两种还原性的电子载体,它们所携带的电子经线粒体内膜上的呼吸链逐级定向传递给O2,本身则被氧化(图6-16)。由于电子传递所产生的质子(H)浓度梯度和电位差,其中所蕴藏的能量被F0F1ATP合酶用来催化ADP磷酸化而合成ATP,这就是氧化磷酸化耦联或氧化磷酸化作用。
呼吸链:
第一条链:NADH+H+、Ⅰ(FMN、FeS)、CoQ、Ⅲ(Cytb、FeS、Cytc1)、Cytc、Ⅳ(CuA、Cyta、Cyta3-CuB)、O2
第二条链:琥珀酸/其他、Ⅱ(FAD、FeS)、CoQ、Ⅲ(Cytb、FeS、Cytc1)、Cytc、Ⅳ(CuA、Cyta、Cyta3-CuB)、O2
琥珀酸→FAD→Fe-S→Q此过程释放的自由能较小,不足以将H+泵出线粒体内膜,因此复合体Ⅱ没有H+泵的功能。根据对相邻电子载体的氧化还原电位和质子数的测定表明,呼吸链中有3个主要的质子由基质转运到膜间腔的部位,即:
NADH→FMN:实际上应该是FeS→Q。复合体Ⅰ泵4H+ 。以下H+转运过程仅绿皮细生明确指出,存疑。而且FMN是L型复合体Ⅰ伸入基质的亲水端的组成部分,其上结合的H怎么可能移动到膜间腔呢?
目前认为复合体Ⅰ的几个亚基在线粒体内膜上形成4个质子通道。电子由FMN通过铁硫簇传递到辅酶Q,会引起质子通道的构象变化,最终完成质子易位。文中“but it is the transfer of electrons to bound Q that drives the pump”提示应该是N2到Q驱动质子泵,N2是硫铁蛋白,所以蓝皮书是错的。 #归档
参考文献:Haapanen O, Sharma V. A modeling and simulation perspective on the mechanism and function of respiratory complex I. Biochim Biophys Acta Bioenerg. 2018 Jul;1859(7):510-523. doi: 10.1016/j.bbabio.2018.04.001. Epub 2018 Apr 13. PMID: 29660310.
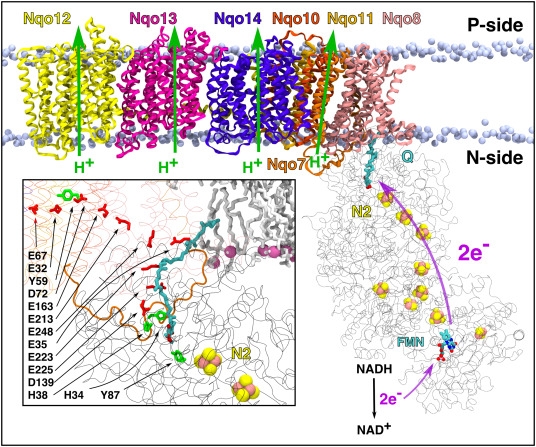
- NADH+H+(基质)+FMN→NAD++FMNH2
- FMNH2+2FeS→2H+(
释放到膜间隙)+(2e-+2FeS) - (2e-+2FeS)+泛醌+2H+(基质)→QH2+2FeS
- 细胞色素b→细胞色素c:复合体Ⅲ泵4H+,至于算不算质子泵?可能也不是不行吧。
- 细胞色素a→O2:复合体Ⅳ确实是质子泵,泵2H+(因为氧化还原电位差的能量只有这么多),2H+被用于合成水。 #归档
参考文献:Proton pumping by cytochrome c oxidase – A 40 year anniversary, Biochimica et Biophysica Acta (BBA) - Bioenergetics, Volume 1859, Issue 9, 2018, Pages 692-698, ISSN 0005-2728, https://doi.org/10.1016/j.bbabio.2018.03.009. (https://www.sciencedirect.com/science/article/pii/S0005272818300422)- Cytc→CuA
- CuA→Cyta
- Cyta→Cyta3-CuB
- Cyta3-CuB+2H++1/2O2→H2O
(很遗憾,下面这个图有很多错误)
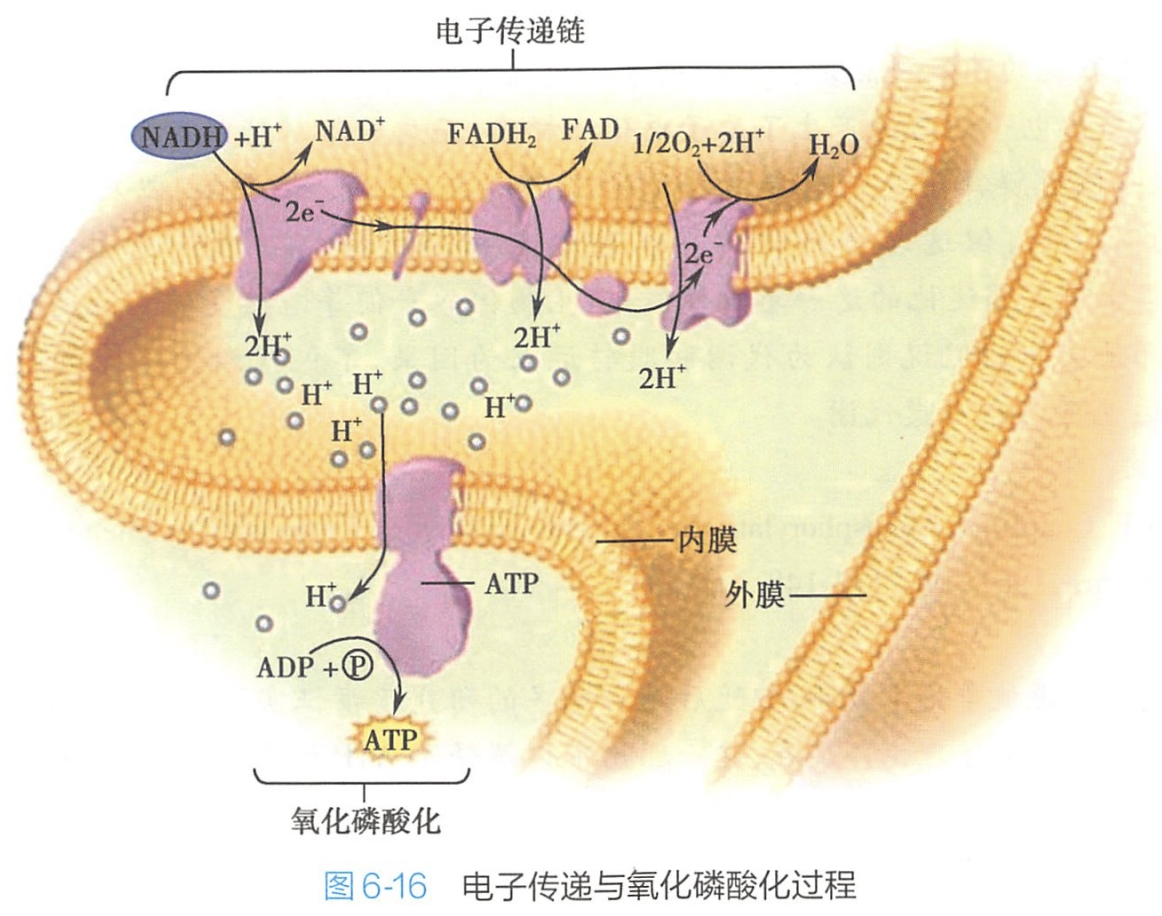
由这些质子在线粒体膜间腔和线粒体基质之间形成的浓度梯度和电位差,足以使2.5分子ADP磷酸化生成2.5分子的ATP。载氢体NADH和FADH2进入呼吸链的部位不同,所形成的ATP也有差异。1分子NADH+H+经过电子传递,释放的能量可以形成2.5分子ATP;而1分子FADH2所释放的能量则能够形成1.5分子ATP。
原理:每得到2个e-泵到膜间隙的H+:4(复合体Ⅰ)+4(复合体Ⅲ)+2(复合体Ⅳ,因为2个H+合成水):4+2=10:6,同时4H+=1ATP(应该是实验所得)
综上所述,葡萄糖完全氧化所释放的能量主要通过两条途径形成ATP:
- 底物水平磷酸化:生成4分子ATP,其中在糖酵解和三羧酸循环中分别生成2分子ATP;
- 氧化磷酸化:生成28个ATP分子。在葡萄糖的氧化过程中,一共产生12对H。其实是显然的,毕竟(CH2O)6+6H2O
- 其中的10对以NAD为载氢体,经氧化磷酸化作用可生成25个ATP分子,
- 2对以FAD为载氢体进入电子传递链,经氧化磷酸化作用可生成3个ATP分子,共产生28个ATP分子。
因此,1分子葡萄糖完全氧化共可生成32分子ATP,其中仅有2分子ATP是在线粒体外通过糖酵解直接形成的。葡萄糖有氧氧化的产能效率大大高出无氧酵解的能量利用效率。
(三)电子传递时H+穿膜形成电化学梯度
化学渗透假说(chemiosmotic coupling hypothesis)。该假说认为氧化磷酸化耦联的基本原理是电子传递中的自由能差造成H+穿膜传递,暂时转变为横跨线粒体内膜的电化学质子梯度(electrochemical proton gradient)。然后,质子顺梯度回流并释放出能量,驱动结合在内膜上的ATP合酶,催化ADP磷酸化合成ATP。这一过程可综合如下:
- NADH或FADH2提供一对电子,经电子传递链,最后为O2所接受;
- 电子传递链同时起H+泵的作用,在传递电子的过程中伴随着H+从线粒体基质到膜间腔的转移;
- 线粒体内膜对H+和OH-具有不可透性,所以随着电子传递过程的进行,H+在膜间腔中积累,造成了内膜两侧的质子浓度差,从而保持了一定的势能差;
- 膜间腔中的H+有顺浓度返回基质的倾向,能借助势能通过ATP酶复合体F0上的质子通道渗透到线粒体基质中,所释放的自由能驱动F0F1ATP合酶合成ATP
化学渗透假说有两个特点,一是需要定向的化学反应;二是突出了膜的结构。
(四)电化学梯度所包含的能量转换成ATP的化学能
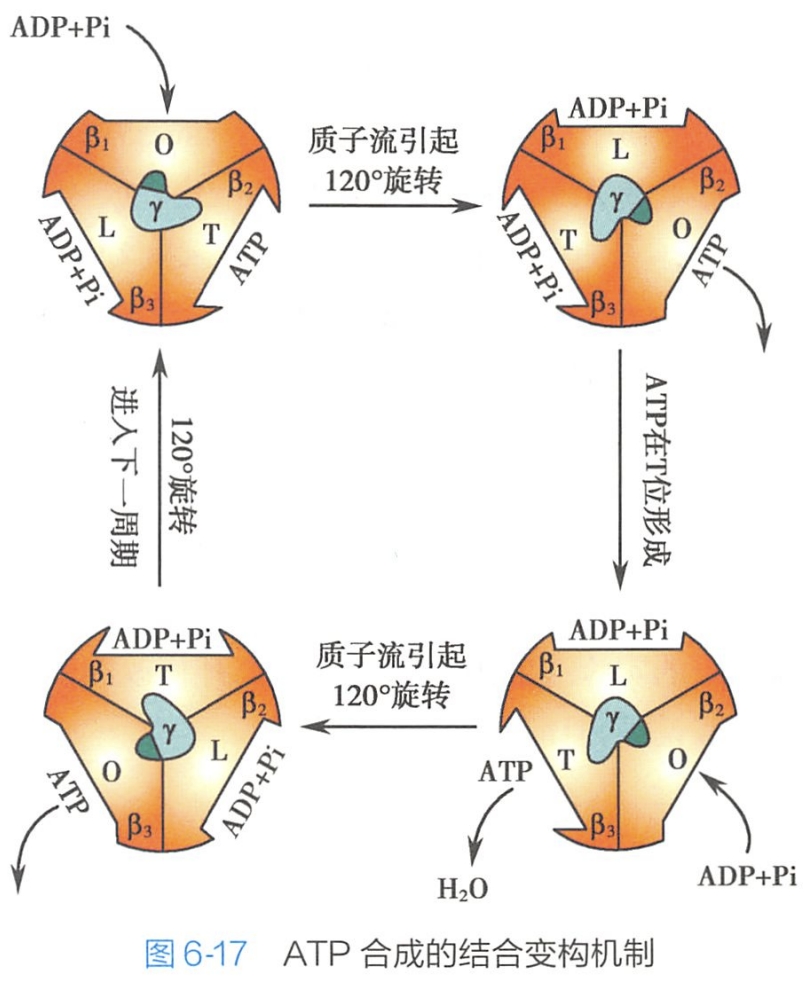
结合变构机制(binding-change mechanism)解释F1因子在ATP合成中的作用过程(图6- 17)。
他认为当质子穿过F1因子( #? 看起来像柄部)的活性部位时可引起F1颗粒的构象变化,导致底物(ADP和 Pi)同活性部位的紧密结合和产物(ATP)的释放。在此模型中,ADP和Pi合成ATP的反应不需要能量,而构象变化时要依赖能量供应。所需的能量一方面是用来使ATP同活性部位的结合由紧密状态变为疏松状态,便于释放ATP;另一方面使ADP和Pi同活性部位的结合由疏松变为紧密以利于ADP同Pi发生反应合成ATP。(注意β没转,只是构象改变。一个循环其实是生成3ATP)
线粒体内膜具有高度不透性,因此这些物质进出线粒体需要依靠专门的结构。线粒体内膜上有一些专一性转运蛋白同这些物质进出线粒体有关,例如其中的一种为腺苷酸转移酶能利用内膜内外H梯度差把ADP和Pi运进线粒体基质,而把ATP输往线粒体外。 #生化